Sensitive environments
Sensitive environments were identified using the MarLIN method developed by the United Kingdom’s Marine Life Information Network.

Sensitivity is dependent on the tolerance of a species or habitat to damage from an external factor, and the time taken for its subsequent recovery from damage sustained as a result of an external factor. Assessments of tolerance takes rarity into account, as the more rare a habitat is, the more an external factor is likely to damage a significant proportion of the habitat, and therefore it will have a lower tolerance rating.
If a habitat or species is very adversely affected by an external factor arising from human activities or natural events (killed/destroyed) and is expected to recover over a very long period of time, (10-20 years) then it would be considered to be highly sensitive. Similarly, if a habitat or species is adversely affected by an external factor arising from human activities or natural events (damaged) but is expected to recover in a short period of time (1-5 years) then it would be considered to be of low sensitivity.
Sensitive environments are identified in Schedule 6 of the Regulations, which also sets out indicators for determining if a sensitive environment is present. They are described in more detail in a NIWA publication ‘Sensitive marine benthic habitats defined’. However, a lack of information means that the definitions are to be regarded as “preliminary and work-in-progress”.
The sensitive environments are:
Stony coral thickets or reefs

Coldwater corals are most commonly found between 200 m and 2000 m water depth and at temperatures between 4°C and 12°C. Coldwater corals are slow growing and long lived. They require a hard substrate for attachment as well as high nutrient and food supply for growth, currents or mixing to deliver the food and nutrients and low sedimentation rates to allow efficient feeding and to avoid physical burial. There are more than 10,000 described species of coldwater corals worldwide, distributed among the stony corals, soft corals, black corals and hydrocorals.
Coldwater corals can provide habitat for some species of fish and invertebrates. The most complex habitat is provided by the stony coral species which produce three dimensional matrix colonies that can coalesce to form ‘reef’, ‘mound’ or ‘thicket’ structures. These structures can extend over kilometres in length and up to 35 m in height. The location of coldwater coral reefs is not well documented in the southern hemisphere. Small patch reefs or thickets (up to 600 m long, 20 m wide, and 3 m high) have been directly observed on many seamounts around New Zealand. Dispersed thickets form where a substrate for attachment is provided by isolated rocks or stones (such as the phosphorite nodules that dominate the crest of the Chatham Rise).
Coldwater corals are fragile and the biodiversity associated with these coral structures can be high. As such these habitats are considered to be ‘vulnerable marine ecosystems’ that require protection from the impacts of deep-water fishing, drilling and mining.
Xenophyophore beds

Xenophyophores are very large, single celled protozoans. They live on the seabed and from complex clumps up to 25 cm in diameter made up of mineral grains, sponge spicule fragments and organic debris. Most of any xenophyophore is dead matter - living plasma makes up less than 5% of the clump volume. They may be easily mistaken for broken and decayed parts of other animals or for inorganic concrements.
Xenophyophores are fact growing. They feed on fine particles, such as bacteria, from the seabed or the water column. Seven species have been recorded in New Zealand, three of which are endemic. It is expected that more species will be recorded.
Xenophyophores are associated with higher infaunal densities. Xenophyophores are particularly abundant below areas of high surface productivity. They have been located on the eastern, northern and western continental slopes of New Zealand and on the Chatham Rise.
Bryozoan thickets

Bryozoans (commonly called moss animals) are suspension feeding organisms that live in colonies, either on the benthos or are epibiotic on algae, seagrass, and animals. The individuals that comprise a colony are called zooids and are less than a millimetre in size. Colonies have varying growth forms and their size varies. They can get as large as 1000 mm across. Bryozoans resemble corals but are unrelated.
Bryozoans contribute significantly to habitat complexity. Other frame building taxa (molluscs, sponges, corals, etc) are an important characteristic of bryozoan habitat and may facilitate the growth of bryozoans by providing a stable substrate on which they can grow or by ‘welding’ branches together. Bryozoans provide a larger surface area than the area of sea floor they occupy. Thickets alter local physical processes such as current speed. Bryozoans also trap sediments within their structures.
The term ‘thicket’ refers to frame-building species which dominate square metres of sea floor.
Habitat-forming bryozoans are particularly abundant and diverse in New Zealand. 27 species provide habitat over hundreds of square kilometres of sea floor. Important species include Cinctipora elegans, Celleporaria agglutinans, and Hippomenella vellicata.
Calcareous tube worm thickets

New Zealand has a number of tube worm species that secrete tubes of calcium carbonate. They are most common in coastal waters, but occur at all depths. They are found from Taranaki south to Stewart Island.
Tube worm species may form clusters that result in the growth of intertwined calcareous tubes which form mounds or patch reefs. Mounds may occur on soft and hard seabed but generally need some hard substrate on which to establish. Mounds are likely to be rare in New Zealand’s EEZ.
Chaetopteridae worm fields

A number of tubeworm species reach sufficient sizes and/or densities to provide biogenic habitat for other species. Low relief worm tube meadows are widespread in many areas. In New Zealand, virtually nothing is known about the potential role of tube-worms in forming biogenic habitat for other species.
One species that forms meadows is Phyllochaetopterus socialis. It lives in a thin wiry tubes 8-10 cm in length which protrude from soft seabed sediments. It can be found as isolated individuals within mixed epifaunal assemblages as well as dense mono-specific meadows which are tens of kilometres in size. This species is also found in waters off Europe, eastern North America, and Australia. Fields are known off the east coast of the South Island though they may also occur around the North Island.
Sea pen field

Sea pens are colonial marine cnidarians (other cnidarians include sea anemones, corals, and jellyfish). Sea pens anchor in the sediment with a root-like bulbous peduncle and carry a feeding polyps on a flexible erect stalk. They occur on soft sediments (fine gravels, soft sand, mud or abyssal ooze) in the deep waters where turbulence is unlikely to dislodge them and where there is a current to ensure a flow of plankton across their feeding polyps. They have mostly been observed as isolated individuals and in a few places in densities of up to 6 per m2 although this may reflect the limited exploration of New Zealand’s EEZ.There are currently 31 species of sea pens known from New Zealand waters, although 19 of those have not been formally described. Fauna associated with New Zealand sea pens have not yet been described.
Rhodolith beds
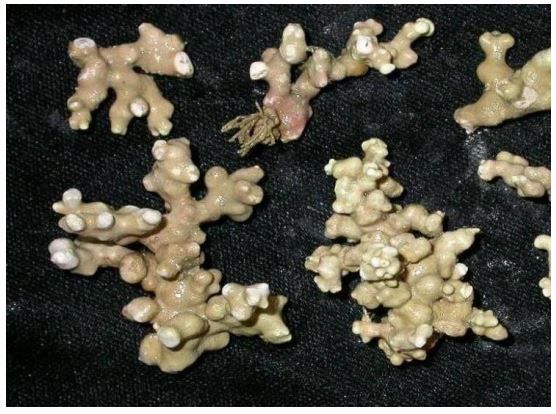
Rhodoliths are free-living calcified red algae which form structurally and functionally complex habitats. They are fragile and slow growing (0.05 – 2 mm/year). Very little information exists about the location, extent or ecosystem functioning of rhodolith beds in New Zealand. It is likely that they occur in the EEZ at localities characterised by strong currents within the photic zone (layer that receives sunlight) to depths of 200m depending on water clarity, particularly around the margins of reefs or elevated banks.
Rhodolith beds feature high benthic biodiversity supporting many rare and unusual species. Productive fisheries often coincide with rhodolith beds, which have been identified as important nursery areas for commercial species. Internationally rhodolith beds have been identified as critically important biodiversity hotspots, harbouring high diversity and abundance of marine animals and algae in comparison with surrounding habitats.
Rhodoliths are at risk from the impacts of a range of human activities including physical disruption (trawling, dredging, anchoring), reduction in water quality (offshore dumping), alterations to water movement (marine engineering) and aquaculture installations (shellfish rafts and lines, fish cages). Rhodoliths, like other calcified macroalgae, will be impacted by acidification of the oceans resulting from global climate change.
Sponge gardens
Sponges are sedentary, filter-feeding metazoans that utilise a single layer of flagellated cells to direct a water current through their bodies for the purposes of feeding and excretion. They may occur on hard or soft substrate in many subtidal environments including shallow coastal rocky reefs, seamounts, hydrothermal vent systems, and oceanic ridges. Well over 500 sponge species have been formally described from New Zealand waters but there are many more known with new discoveries every year.
Certain areas in New Zealand’s EEZ have been identified as ‘hotspots’, areas that are considered significant in terms of sponge biodiversity. The dominant New Zealand sponge biodiversity hotspot is the northern New Zealand region that includes Three Kings, North Cape, Spirits Bay and offshore carbonate bryomol gravel banks.
Sponge gardens are defined and recognised by their high sponge density, but may also be characterised by morphological diversity, the large size and abundance of sponge individuals, percentage cover, and the uniformity or mixed nature of the distribution of the species.


Beds of large bivalve molluscs
Bivalve molluscs are shellfish with two hinged shells (such as mussels, cockles, oysters, pipi). 07024d69-3391-4c7e-921f-58aa7d0df4c1
The list of bed-forming species is too numerous to list exhaustively.
Bivalve beds create biogenic structure in what may be an otherwise ‘featureless’ habitat. In addition, their shells provide a substrate for settlement by organisms such as sponges and bryozoans, and shelter for mobile invertebrates and fishes. These aggregations modify the habitat considerably from that surrounding it, and consequently influence the composition of the associated community.
Bivalve beds create habitat heterogeneity and have an important role in ecosystem functioning. Infaunal bivalves influence biogeochemical processes such as regeneration of sediment-associated nutrients, processing these nutrients and thus making them available for water column primary production. Sediment derived nutrients are considered a major contributor to continental shelf production.
Beds of large bivalves are generally located at depths of less than 250 m and are well represented off Northland, off the west coast of the North Island and off south-eastern and southern-most New Zealand.


Macro-algae beds

Macro-algae range from small foliose (leaf-like shape) brown, red and green algae to large brown algae or kelp. Beds of macro-algae occur on hard rocky substrates within the photic zone (layer that receives sunlight) to depths of 200m depending on water clarity. Within the EEZ rocky reefs in the photic zone are very rare and thus beds of macro-algae are likely to be equally rare. They have, however, been recorded in the EEZ on the Mernoo Bank east of Banks Peninsula, the Ranfurly Bank off East Cape and at least one seamount on the Kermadec Ridge.
Kelps beds are recognised as key contributors to reef ecosystems through the energy captured via photosynthesis, the provision of highly structured three dimensional habitats critical for other species, and also through the fixed carbon retained within, and exported from, kelp forests.
Brachiopod beds

Brachiopods (commonly called lamp shells) are small (adult shells are typically 5-50 mm in length), bilaterally symmetrical, filter feeders, superficially resembling bivalve molluscs. They are generally anchored to a hard substrate by a muscular stalk. New Zealand has 38 species, 18 species of which are endemic to New Zealand. Some species form dense beds 2 or 3 layers deep and up to 1000 individuals per square metre. In some areas dead brachiopod shells contribute to habitat complexity and provide abundant interstices for small invertebrates and fish.
Brachiopods occur throughout New Zealand at all depths. Known diverse or abundant brachiopod assemblages occur off the Three Kings Islands, off Ranfurly Bank (East Cape), parts of the Chatham Rise where rare species are associated with coral thickets, and areas of the Campbell and Bounty Plateaux.
Deep-sea hydrothermal vents

Hydrothermal vents occur where cold seawater percolates down through the seabed, is heated by geothermal energy, become buoyant and rises to the seabed, and dissolves metals and sulphides from the surrounding rocks. Venting can occur from point sources, chimneys (made from precipitated minerals) or crack and fissures in the seabed, or percolate in a diffuse fashion through sands or muds. Hydrothermal vents are associated with the subduction zone of the Pacific plant under the Australian plate to the north of New Zealand.
Hydrothermal vents provide habitat for unique communities of organisms which are dependent on the sulphide-rich vent fluids that support chemosynthetic bacteria (primary producers which use chemicals, instead of light, to create energy) at the base of the food web. Vent specialists can only exist in close proximity to the active venting and this vent community is surrounded by a distinct non-vent community as the periphery of the vent site. The non-vent community is comprised of organisms found elsewhere in the region but often at greater densities due to enhanced food supply. The composition of the communities present at hydrothermal vents is influence by the temperature of venting, distance from the source of venting, and evolutionary and geographic factors.
Seabed communities found at hydrothermal vents are considered to be sensitive to physical disturbance. Despite vent species being adapted to periodic natural disturbance (venting may be periodically ‘witched off’) levels of endism in vent communities suggest impacts to these habitats may have significant effects, including species extinction. Some vent species in New Zealand waters are recognised by DOC’s Threat Classification System, including Vulcanolepis osheai.
Methane or cold seeps

Cold seeps occur where methane-rich fluids escape into the water column from underlying sediments. They are usually associated with areas where gas hydrates (methane frozen within an ice matrix) occur within the sediments, generally in water depths of at least 500 m. Methane seep sites have been identified at many sites on the Hikurangi Margin along the east coast of the North Island. Most of these sites support live communities of obligate seep-associated fauna. Clusters of active methane seeps have also been confirmed within five areas of the east coast of the lower North Island. It is highly likely that more sites are yet to be discovered around New Zealand.
Cold seeps typically support communities dominate by chemoautotrophic benthic organisms which depend on symbioses with chemosyntheic bacteria that generate energy from methane (and other compounds). Seep fauna typically include large tube worms, vesicomyid clams, and bathymodiolin mussels. Sepp fauna may also include other bivalves, gastropods, sponges, shrimp, crustaceans and polychaetes. Much of the seep-associated fauna from the Hikurangi Marin appears to be new to science or endemic to New Zealand seeps, suggesting the region may be a new biogeographic province for cold-seep fauna.
Last updated at 4:43PM on December 19, 2014